RNA甲基化m6A-seq 研究一本通将从m6A是什么,m6A甲基化酶的种类及功能,m6A检测方法汇总,m6A课题设计思路,m6A论文发表梯度推荐,m6A后期验证工具大全到经典案例解析等方面为大家揭开m6A的神秘面纱,每天5分钟,就能从m6A小白入门到高阶提高哦~
往期内容回顾:
m6A甲基化整体研究思路:m6A相关SCI论文发表要求分类汇编
今天的文章将围绕RNA甲基化m6A后期验证工具展开,详细介绍了几类高频的验证实验,一起来学习吧~
前言做完测序后需要对分析结果中感兴趣的内容进行生物学功能验证,一个完整的功能验证将历经分子细胞→细胞功能→动物实验等形成完整的实验思路。其中分子细胞包括RNA-RNA互作验证(如miRNA与UTR互作验证)等。细胞功能包括敲低/敲除/过表达、蛋白细胞定位、蛋白与蛋白互作、蛋白与RNA/DNA互作、细胞表型实验(迁移侵袭、划痕、增殖、凋亡、周期)等。最后的动物实验主要指动物模型实验,如小鼠、斑马鱼等。除了qPCR外,接下来的内容中会选择其中几个高频的验证实验做介绍。
RNA-RNA/DNA互作验证提起RNA或DNA分子互作验证,就不得不提一下双荧光素酶报告基因检测系统。荧光素酶已成为科研中运用最广泛的报告基因之一,其具有检测灵敏度高、结果重复性好、信噪比较高和易于检测等特点。很多实验室使用的荧光素酶报告基因载体通常采用了萤火虫荧光素酶/海参荧光素酶双报告基因系统,其中萤火虫荧光素酶作为检测报告基因,而海参荧光素酶则作为内参报告基因。

因为哺乳动物细胞中不含内源性荧光素酶,一旦转录完成立刻就生成功能性萤光素酶。单报告基因实验往往会受到各种实验条件的影响,而双报告基因则通过共转染的“对照”作为内参为实验提供一个基准线,从而可以在最大程度上减小细胞活性和转染效率等外在因素对实验的影响,使得数据结果更为可信。
Dual-Luciferase 双荧光素酶报告基因检测系统在细胞中同时表达海参荧光酶。两者没有种源同源性并且对应不同的反应底物,故而没有交叉干扰。得益于超强的光信号和超高的信噪比,本系统被广泛用于启动子活性验证、甲基化酶识别位点验证、miRNA靶基因验证、circRNA 与miRNA互作等实验。具体核酸互作的相关protocol详见:https://link.springer.com/protocol/10.1007/978-1-4939-1708-2_4。
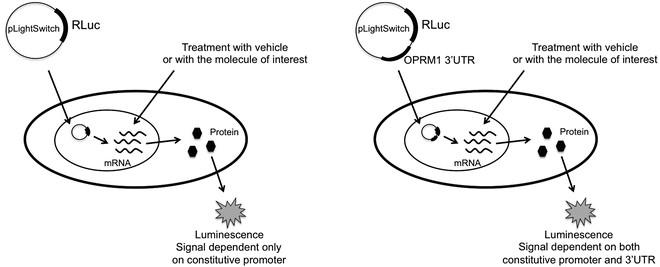
由于miRNA主要通过作用于靶基因的3'UTR起作用,可以将目的基因3'UTR 区域构建至载体中的报告基因luciferase后面,通过比较过表达或者干扰miRNA后,报告基因表达的改变(监测荧光素酶活性变化)可以定量反映miRNA对目的基因的抑制作用;结合定点突变等方法确定miRNA 与靶基因3'UTR 的作用位点。
转录因子是一种具有特殊结构、行使调控基因表达功能的蛋白质分子,也称为反式作用因子。某些转录因子仅与其靶启动子中的特异序列结合,这些特异性的序列被称为顺式因子,转录因子的DNA结合域和顺式因子实现共价结合,从而对基因的表达起抑制或增强的作用。
荧光素酶报告实验是检测这类转录因子和靶启动子中的特异顺序结合的重要手段,其原理简述如下: 第一,用靶启动子的特定片段替换报告基因质粒(如pG L3-basic)的启动子区。第二,将要检测的转录因子表达质粒与报告基因质粒共转。如果该转录因子能移激活靶启动子,则荧光素酶就会表达,且其表达量与转录因子的作用强度成正比。第三,加入特定的底物,荧光素酶与底物反应,通过产生荧光的强度可以对荧光素酶的活性进行定量,进而判断该转录因子能否与靶启动子片段发生作用(及其作用强度)。
敲低/敲除/过表达
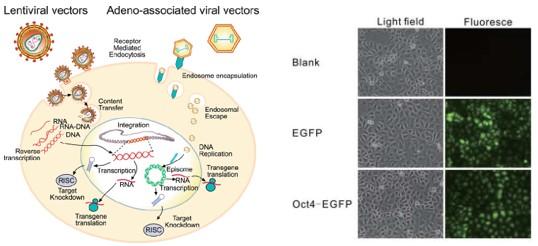
通常研究一类基因或miRNA、lncRNA等在细胞中行使什么样的功能,需要在细胞中对该基因进行敲低(knockdown)、敲除(knockout)或过表达(overexpression)。这就需要将这些干扰质粒或过表达质粒在慢病毒(Lentivirus)、腺相关病毒(adeno-associated virus)、腺病毒(adenovirus)等入侵载体的帮助下,将质粒转染进细胞体内。
病毒是基因操作可视化的杰出载体。通常慢病毒能快速介导基因敲低/敲除/过表达,可做成细胞稳定株进行长期实验。因为外源基因如果不整合进宿主DNA,只能维持几天的高表达。传统的稳定株构建方法需要通过外源基因的瞬时转染后对靶细胞进行筛选。而慢病毒是一种RNA病毒,携带的外源基因在病毒侵染细胞后需要逆转录为DNA,再整合到宿主细胞中后才能表达。这种方法克服了传统方法的弊端,可在短时间内获得大量稳定细胞株。
腺病毒为双链DNA病毒,与慢病毒不同的是其并不会与宿主DNA发生整合。细胞被侵染后在1-3天内表达,并能够持续1-2周,扩散能力强,并且免疫原性较强。
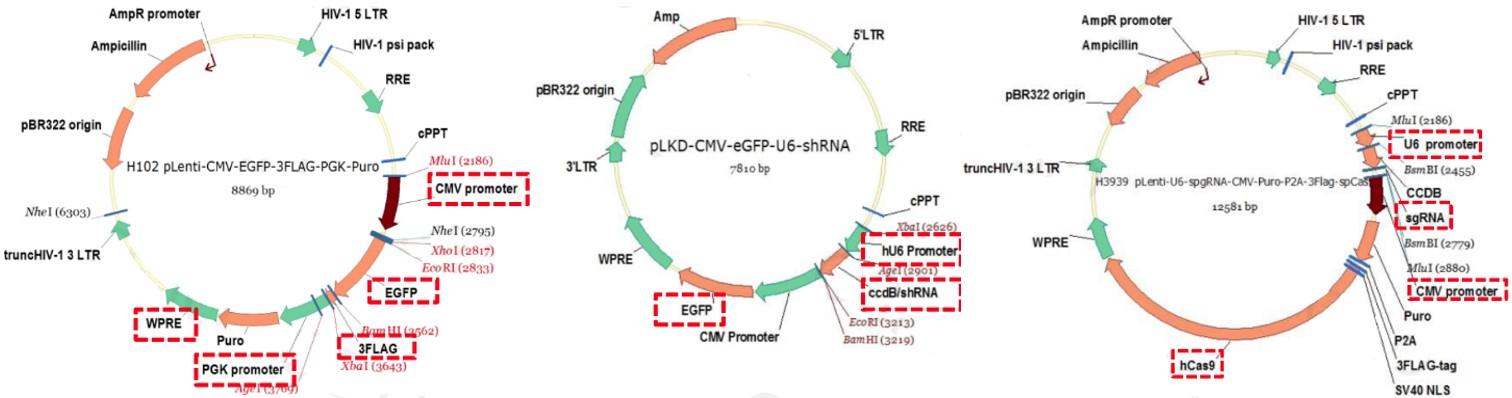
上图中的三个质粒载体示例:分别在基因层面上实现过表达、敲低和敲除的功能。
通常质粒载体需要一个最基本的基因表达结构,包括启动子、终止子基因等。其中CMV、U6都是很常见的启动子,后面能放入各种需要表达的基因。另外EGFP是用来发光的,cas9是CRISPR基因敲除系统中很重要的一种酶,配合gRNA就能对目标区域的基因进行敲除。
今天的内容讲完啦,大家记得收藏好慢慢学习哦~
以上后期验证服务联川也提供的哦~
下期预告:RNA 甲基化 m6A 后期验证工具之细胞蛋白定位/免疫荧光实验及蛋白与蛋白互作(免疫共沉淀和 GST pull down)
相关阅读推荐:
m6A甲基化整体研究思路:m6A相关SCI论文发表要求分类汇编
史上最详细RNA甲基化套路解读 | m6A调节果蝇性别分化和神经功能
Nature:m6A调控miRNA初级体识别加工 | RNA甲基化套路解读
RNA甲基化套路解读三:m6A识别酶HNRNPA2B1介导RNA加工